
Hearing, the cochlea, the frequency domain and Fourier’s series
In recent weeks, we have seen repeated attempts to suggest that Mathematics is essentially a mind game we make up as an aspect of culture. There has been a very strong resistance to the idea that there are intelligible manifestations of structure and quantity embedded in the fabric of the world (and indeed in that of any possible world). And when test cases have been put on the table, they have been consistently brushed aside as cases where our mathematical modelling has been applied; that is it’s all in our heads.
So, it is appropriate to put on the table a test case that is quite literally in our heads, hearing and particularly how the cochlea works. Video:
We see here how there is a frequency domain transformation that makes use of the mechanical properties of the inner ear. That is, our hearing moves from the time to the frequency domain, sensing pitch; also, subtle timing differences between sound arrivals at our right and left ears help us to locate sound sources in the space around us. As was noted in a comment in the Fourier thread:
_____________
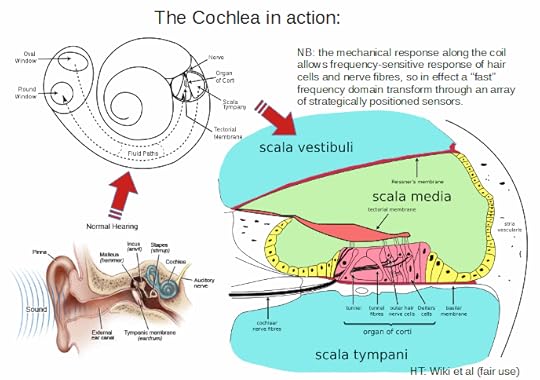
KF, 4: >>On how hearing creates a frequency domain transform of sound inputs, driving the onward processing, Wiki is a handy reference:
The stapes (stirrup) ossicle bone of the middle ear
transmits vibrations to the fenestra ovalis (oval window) on the outside
of the cochlea, which vibrates the perilymph in the vestibular duct
(upper chamber of the cochlea). The ossicles are essential for efficient
coupling of sound waves into the cochlea, since the cochlea environment
is a fluid–membrane system, and it takes more pressure to move sound
through fluid–membrane waves than it does through air; a pressure
increase is achieved by the area ratio of the tympanic membrane to the
oval window, resulting in a pressure gain of about 20× from the original
sound wave pressure in air. This gain is a form of impedance matching –
to match the soundwave travelling through air to that travelling in the
fluid–membrane system . . . .The perilymph in the vestibular duct and the endolymph in the
cochlear duct act mechanically as a single duct, being kept apart only
by the very thin Reissner’s membrane. The vibrations of the endolymph in the cochlear duct displace the basilar membrane in a pattern that peaks a distance from the oval window depending upon the soundwave frequency.
The organ of Corti vibrates due to outer hair cells further amplifying
these vibrations. Inner hair cells are then displaced by the vibrations
in the fluid, and depolarise by an influx of K+ via their
tip-link-connected channels, and send their signals via neurotransmitter
to the primary auditory neurons of the spiral ganglion.The hair cells in the organ of Corti are tuned to certain sound frequencies by way of their location in the cochlea,
due to the degree of stiffness in the basilar membrane.[3] This
stiffness is due to, among other things, the thickness and width of the
basilar membrane,[4] which along the length of the cochlea is stiffest
nearest its beginning at the oval window, where the stapes introduces
the vibrations coming from the eardrum. Since its stiffness is high
there, it allows only high-frequency vibrations to move the basilar
membrane, and thus the hair cells. The farther a wave travels towards
the cochlea’s apex (the helicotrema), the less stiff the basilar
membrane is; thus lower frequencies travel down the tube, and the
less-stiff membrane is moved most easily by them where the reduced
stiffness allows: that is, as the basilar membrane gets less and less
stiff, waves slow down and it responds better to lower frequencies.
In addition, in mammals, the cochlea is coiled, which has been shown to
enhance low-frequency vibrations as they travel through the fluid-filled
coil.[5] This spatial arrangement of sound reception is referred to as
tonotopy . . . . Not only does the cochlea “receive” sound, it generates
and amplifies sound when it is healthy. Where the organism needs a
mechanism to hear very faint sounds, the cochlea amplifies by the
reverse transduction of the OHCs, converting electrical signals back to
mechanical in a positive-feedback configuration. The OHCs have a protein
motor called prestin on their outer membranes; it generates additional
movement that couples back to the fluid–membrane wave. This “active
amplifier” is essential in the ear’s ability to amplify weak
sounds.[6][7]The active amplifier also leads to the phenomenon of soundwave
vibrations being emitted from the cochlea back into the ear canal
through the middle ear (otoacoustic emissions) . . . .Otoacoustic emissions are due to a wave exiting the cochlea via the
oval window, and propagating back through the middle ear to the eardrum,
and out the ear canal, where it can be picked up by a microphone.
Otoacoustic emissions are important in some types of tests for hearing
impairment, since they are present when the cochlea is working well, and
less so when it is suffering from loss of OHC activity . . . .The coiled form of cochlea is unique to mammals. In birds and in
other non-mammalian vertebrates, the compartment containing the sensory
cells for hearing is occasionally also called “cochlea,” despite not
being coiled up. Instead, it forms a blind-ended tube, also called the
cochlear duct. This difference apparently evolved in parallel with the
differences in frequency range of hearing between mammals and
non-mammalian vertebrates. The superior frequency range in mammals is
partly due to their unique mechanism of pre-amplification of sound by
active cell-body vibrations of outer hair cells. Frequency resolution
is, however, not better in mammals than in most lizards and birds, but
the upper frequency limit is – sometimes much – higher. Most bird
species do not hear above 4–5 kHz, the currently known maximum being ~
11 kHz in the barn owl. Some marine mammals hear up to 200 kHz. A long
coiled compartment, rather than a short and straight one, provides more
space for additional octaves of hearing range, and has made possible
some of the highly derived behaviors involving mammalian hearing . . .
In short, sinusoidal frequency domain decomposition of sound waves is a key mechanical phenomenon exploited by our hearing system, leading to in effect a frequency domain transformation of the temporal pattern of compressions and rarefactions that we term sound. This is of course closely related to the patterns we explored and discovered using Fourier power series and integral analysis of oscillations and transient pulses.
Where, on the mechanical side, harmonic motion is tied to elastic and
inertial behaviour. Which in turn is directly connected to a rotating
vector analysis — leading straight to the complex exponential analysis
that draws out the full power of complex numbers, form Z = R*e^i*wt, w
being circular frequency 2* pi*f (in radians per second), f the cycle
per second frequency. All of this ties back to the fundamental frequency
cycles and integer-multiple frequency harmonic epicycles in the OP
above.
Again, Mathematical study turns out to reflect quantities, structures
and linked phenomena which are embedded in the fabric of our world.
KF
PS: Notice, not a few design subtleties?
PPS: The vocal tract, in effect a wind instrument, also exploits
fundamentals and harmonics to create auditory, frequency-based patterns
as well as transients.>>
______________
We see here yet another case where structure and quantity are embedded in the natural world and are exploited in the design of our bodily organs; here, those for hearing. Thus, literally in our heads. END
Copyright © 2019 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.
Plugin by Taragana


Michael J. Behe's Blog

- Michael J. Behe's profile
- 219 followers

