
Michael J. Behe's Blog, page 27
November 9, 2022
At Science Daily: Early planetary migration can explain missing planets
Computer simulations by Rice University scientists and their collaborators are the first to integrate a model of planet formation and evolution that explains two puzzling observations of exoplanets orbiting distant stars: the rarity of worlds about 1.8 times larger than Earth and the near-identical size of adjacent planets in hundreds of planetary systems.
A new model that accounts for the interplay of forces acting on newborn planets can explain two puzzling observations that have cropped up repeatedly among the more than 3,800 planetary systems cataloged to date.
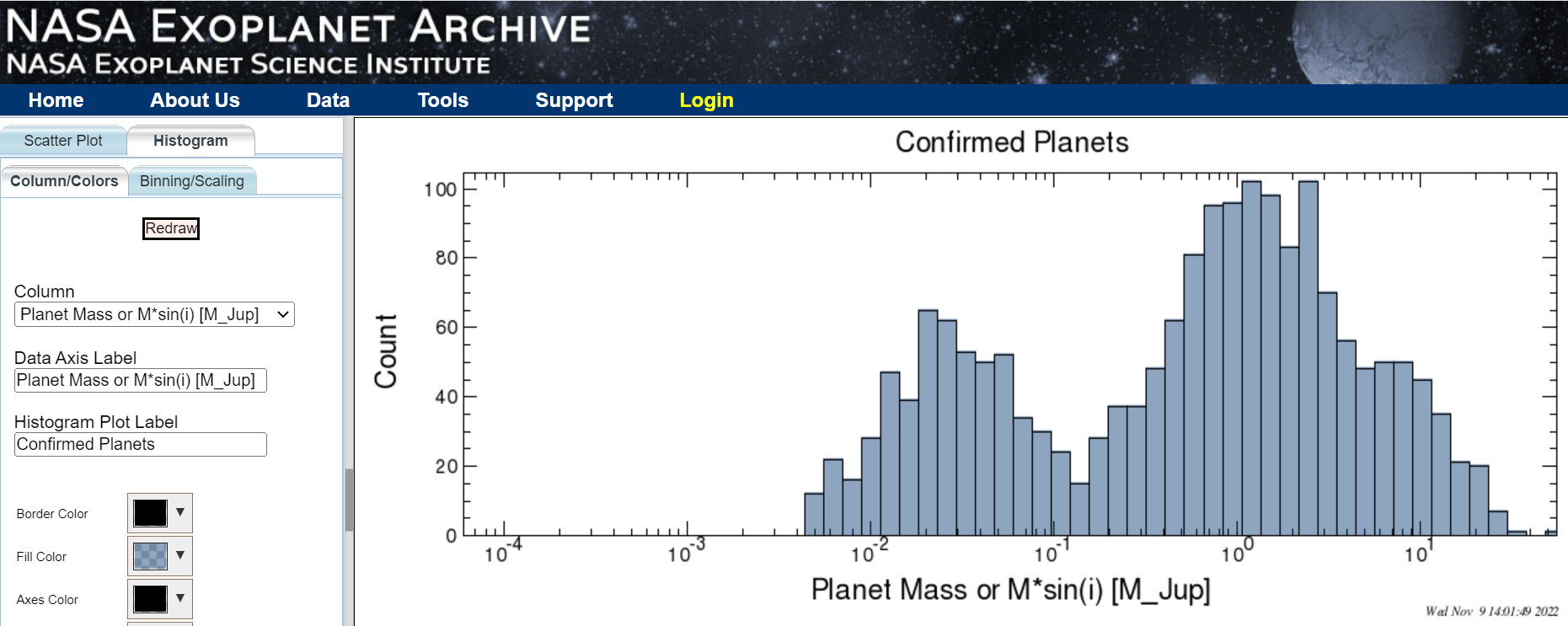 Plot of distribution of confirmed exoplanet masses. Note that Earth’s mass is about 0.003 Jupiter masses, or just to the left of the left-most column in the histogram.
Plot of distribution of confirmed exoplanet masses. Note that Earth’s mass is about 0.003 Jupiter masses, or just to the left of the left-most column in the histogram.One puzzle known as the “radius valley” refers to the rarity of exoplanets with a radius about 1.8 times that of Earth. NASA’s Kepler spacecraft observed planets of this size about 2-3 times less frequently than it observed super-Earths with radii about 1.4 times that of Earth and mini-Neptunes with radii about 2.5 times Earth’s. The second mystery, known as “peas in a pod,” refers to neighboring planets of similar size that have been found in hundreds of planetary systems. Those include TRAPPIST-1 and Kepler-223, which also feature planetary orbits of near-musical harmony.
“I believe we are the first to explain the radius valley using a model of planet formation and dynamical evolution that self-consistently accounts for multiple constraints of observations,” said Rice University’s André Izidoro, corresponding author of a study published this week in Astrophysical Journal Letters. “We’re also able to show that a planet-formation model incorporating giant impacts is consistent with the peas-in-a-pod feature of exoplanets.”
Izidoro, a Welch Postdoctoral Fellow at Rice’s NASA-fundedCLEVER Planets project, and co-authors used a supercomputer to simulate the first 50 million years of the development of planetary systems using a planetary migration model. In the model, protoplanetary disks of gas and dust that give rise to young planets also interact with them, pulling them closer to their parent stars and locking them in resonant orbital chains. The chains are broken within a few million years, when the disappearance of the protoplanetary disk causes orbital instabilities that lead two or more planets to slam into one another.
“The migration of young planets towards their host stars creates overcrowding and frequently results in cataclysmic collisions that strip planets of their hydrogen-rich atmospheres,” Izidoro said. “That means giant impacts, like the one that formed our moon, are probably a generic outcome of planet formation.”
The research suggests planets come in two “flavors,” super-Earths that are dry, rocky and 50% larger than Earth, and mini-Neptunes that are rich in water ice and about 2.5 times larger than Earth. Izidoro said new observations seem to support the results, which conflict with the traditional view that both super-Earths and mini-Neptunes are exclusively dry and rocky worlds.
Based on their findings, the researchers made predictions that can be tested by NASA’s James Webb Space Telescope. They suggest, for instance, that a fraction of planets about twice Earth’s size will both retain their primordial hydrogen-rich atmosphere and be rich in water.
Full article at Science Daily.
Planetary sizes, masses, and orbital distances from their host stars are found to not be uniformly distributed. Most exoplanets discovered so far are much more massive than Earth, larger in radius than Earth, and orbit much closer to their host star than Earth. None of these deviations from Earth-likeness would be advantageous for advanced life.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana


At Evolution News: William Dembski Offers an Updated Edition of an Intelligent Design Classic
 Photo: William Dembski, via Discovery Institute.
Photo: William Dembski, via Discovery Institute.
On a new episode of ID the Future, mathematician and philosopher William Dembski talks with host Eric Anderson about a revised and updated edition of Dembski’s pioneering 1998 Cambridge University Press book, The Design Inference. Dembski says he stands by that work and his early contributions to intelligent design theory, but adds that he has learned a lot more in the intervening years, particularly from his work with Robert J. Marks and Winston Ewert at the Evolutionary Informatics Lab. Lessons from that and other research, Dembski explains, will enrich the new edition. What light do these design-detecting methods shed on modern evolutionary theory? To find out, download the podcast or listen to it here.
Evolution News
[William Dembski begins to discuss his updated edition of The Design Inference at about 7 minutes into the podcast.]
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
Elaborated Tunnel Architectures in Enzyme Systems point to a designed setup
Elaborated Tunnel Architectures in Enzyme Systems point to a designed setup
RNA and DNA belong to the four basic building blocks of life. They are complex macromolecules made of three constituents: the base, the backbone, which is the ribose five-carbon sugar, and phosphate, the moiety which permits DNA polymerization and catenation of monomers, to become polymers. The nucleobases are divided into pyrimidine and purines. These bases must be made in complex biosynthesis pathways in the cell, requiring several molecular machines, and enzymes, that perform the gradual, stepwise operations to yield the nucleobases, which, in the end, are handed over for further processing. Pyrimidines, one of the two classes, require 7 enzymes, of which Carbamoyl phosphate synthase II is the first in the production line.
In bacteria, a single enzyme supplies carbamoyl phosphate for the synthesis of arginine and pyrimidines. The bacterial enzyme has three separate active sites, spaced along a tunnel nearly 100 Å long. Bacterial carbamoyl phosphate synthetase provides a vivid illustration of the channeling of unstable reaction intermediates between active sites. This reaction consumes two molecules of ATP: One provides a phosphate group and the other energizes the reaction. The need for this channel exists to efficiently translocate reactive gaseous molecules that can either be toxic to the cell or are reactive intermediates that need to be delivered to complete a coupled reaction.
Comment: Consider that no lifeform exists that does not use DNA and RNA. Therefore, the synthesis of these molecules is a prerequisite for life. The origin of this metabolic pathway can therefore not be explained through evolution. Either it was design or random nonguided fortunate events.
Tunnel Architectures in Enzyme Systems that Transport Gaseous Substrates
Derinkuyu Underground City in Cappadocia, Turkey, is one of the deepest and most fascinating multilevel subterranean cities, excavated in tunnel systems. Specifically constructed, elaborated Air ducts ensure fresh oxygen supply, and the oxygen ratio inside never changes no matter at what level one is in. Such systems are always engineering marvels, and must be precisely calculated, and constructed. Remarkably, some proteins act similarly and exist in molecular biological systems.
Ruchi Anand (2021): Tunnels connect the protein surface to the active site or one active site with the others and serve as conduits for the convenient delivery of molecules. Tunnels transferring small molecules such as N2, CH4, C2H6, O2, CO, NH3, H2, C2H2, NO, and CO2 are termed gaseous tunnels. Conduits that have a surface-accessible connection and can accept gases from the surroundings are named external gaseous (EG) tunnels. Whereas, buried gaseous tunnels that do not emerge to the surface are named internal gaseous (IG) tunnels. In some cases, the tunnels can be performed, permanently visible within the protein structure such that the natural breathing motions in proteins do not alter the tunnel dimensions to the extent that the radius of the gaseous tunnel falls below the minimum threshold diameter, e.g., carbamoyl phosphate synthetase (CPS) has a preformed tunnel. In contrast, it can be transient such that the tunnel diameter is not sufficiently wide enough to allow the incoming molecule to pass through it or certain constrictions in the tunnel block its delivery. This could be either to control the frequency of molecules traveling across or to coordinate and facilitate coupled reaction rates. Another possible scenario of transient tunnel formation is one in which the tunnel is nonexistent in the apo state, and only upon significant conformational change, under appropriate cues, is the tunnel formed. In several cases transient tunnels require intermediate/substrate-induced conformational changes in the tunnel residues to open up for the transport of the incoming molecule, within the respective enzyme. These tunnels undergo enormous fluctuations and switch between open and close states. It is remarkable that the presence of these conduits, which are as long as 20−30 Å and even longer like 96 Å in CPS,6a run inside the protein body, forming pores that serve as highways for transport of these gaseous molecules. In several cases, an added level of tuning into the tunnel architecture is introduced by incorporating gating mechanisms into the EG and IG tunnel architectures.
Gates serve as checkpoints and vary from system to system; some are as simple as an amino acid blocking the path which moves out upon receiving appropriate cues such as the swinging door type in cytidine triphosphate synthase (CTP) and in others more complex arrangement of amino acids come together to form control units such as aperture gates, drawbridge, and shell type gates. These tunnels and their gates are connected via an active communication network that spans between distal centers and hence introduces both conformation and dynamic allostery into the protein systems. It is not uncommon to observe long-distance allosteric networks that can be dynamic in nature and transiently formed via the motion of loop elements, secondary structural rearrangements, or of entire domains.
EXTERNAL GASEOUS (EG) TUNNEL ARCHITECTURES
EG tunnels connect the bulk solvent with the active site of an enzyme. These tunnels are found in several enzymes that accept gaseous substrates to facilitate their delivery to the buried active site. A class of predominant gaseous substrates are alkanes such as methane and ethane gases that are oxidized aerobically or via anaerobic pathways. Recently, the crystal structure of the enzyme that anaerobically oxidizes ethane to ethylCoM from Candidatus Ethanoperedens thermophilum was determined, and named it ethylCoM reductase. The enzyme belongs to the broad methylCoM reductase superfamily, which oxidizes methane. The ethylCoM reductase has a 33 Å tunnel that runs across the length of the protein. Interestingly, the EG tunnel present in ethylCoM reductase has some very unique features. At the end of the tunnel, near the Ni-cofactor F430 active site, there are several residues that are post-translationally modified. Methylated amino acids, such as S-methylcysteine, 3-methylisoleucine, 2(S)-methylglutamine, and N2 -methylhistidine line the tunnel. It is likely that these residues tune the enzyme to select for ethane by creating a very hydrophobic environment and prevent similar-sized hydrophilic molecules such as methanol from reaching the active center. The larger hydrophobic alkanes are selected out via optimization of the tunnel diameter, which is fit to accommodate ethane. Another example of an alkane transporting tunnel exists in soluble methane monooxygenase (sMMO) that performs C− H functionalization by breaking the strongest C−H bond, among saturated hydrocarbons, in methane and aerobically oxidizes it to form methanol. In methanotrophs, these enzymes are tightly regulated, and the complex formation between the two proteins, hydroxylase MMOH and regulatory protein MMOB, is required for function. The EG tunnel formed in this system is very hydrophobic, and the diameter is such that it only allows for smaller gases such as methane and O2 to percolate into the di-Fe cluster harboring active site. In Methylosinus trichosporium OB3b, half of the tunnel is at the interface of the MMOH/MMOB complex, and another half of the tunnel is buried within MMOH, where the oxidation reaction is catalyzed. As an added control feature, the complex has multiple gates to regulate its function. Residues W308 and P215 guard the entrance of the substrate molecules and block the formation of the EG tunnel in the absence of the complex between MMOH and MMOB.
Comment: This demonstrates and exemplifies how in many cases, single monomers have important functions, and changing them through mutations can remove the function of the entirety of the enzyme.
Upon complexation, a conformational change is triggered, and these residues move out of the path, opening the passage for the entire tunnel. When the upper gating residues move upon MMOB/MMOH complex formation, another residue F282 right near the active site also concomitantly undergoes a shift, allowing methane and oxygen to access the di-Fe center. MMOH also has an alternative secondary hydrophilic passage, accessible only when MMOB/MMOH complex dissociates which allows the polar methanol product to be released through it. The gating residues, F282 in the hydrophobic EG tunnel and E240 in the hydrophilic passage, switch between open and close states alternately upon binding/unbinding of MMOB and hence opens one of the two tunnels at a time. This regulates the flow of substrates and products and avoids overoxidation of methanol by releasing it through the hydrophilic passage prior to the entry of substrates in the active site via the hydrophobic EG tunnel.
One of the most common gaseous substrates for which several examples of tunneling enzymes exist is oxygen (O2). It is used in several important oxidation reactions for the generation of essential pathway intermediates and also is a key transport gas in cells. Interestingly in several cases, oxygen is transported to the desired site via molecular tunnels, perhaps to modulate its flow. There are two types of tunnel architectures that are prevalent: first, where there is a main tunnel connected to several subsidiary tunnels, and second, those with fewer tunnels but with stringent gating controls. For instance, soybean lipoxygenase-1 is an example of a multitunnel system that has eight EG tunnels, out of which the one that is formed by hydrophobic residues, such as L496, I553, I547, and V564, has the highest throughput and is identified as the main gaseous tunnel for delivering O2 to the reaction center. It catalyzes the stereospecific peroxidation of linoleic acid via forming a pentadienyl radical intermediate. Under oxygen-deficient conditions, the intermediate escapes from the active site to the bulk and forms four products, i.e., 13S-, 13R-, 9S-, and 9R-hydroperoxy-octadecadienoic acid, in equal distributions. However, under ambient O2 conditions, the EG tunnel delivers O2 efficiently into the active site which has a properly positioned and oriented radical intermediate. Here, O2 is delivered by the EG tunnel such that it stereo- and regiospecifically attacks the radical intermediate to yield 13S-hydroperoxy-octadecadienoic acid as a major product with ∼90% yield. It has also been shown that when the EG tunnel residue L496 is mutated to a bulky tryptophan, it opens up a new gaseous tunnel for O2 delivery, where it attacks at the different side of the pentadienyl intermediate, preferring the formation of 9S- and 9Rproducts. This example showed the importance of the gaseous tunnel in determining the stereo- and regiospecificity for product formation
INTERNAL GASEOUS (IG) TUNNEL ARCHITECTURES
While the EG tunnels transport gases and have pores that are accessible to the surface, there is another class of tunnels formed within the core of the enzyme system, buried in the body of the protein, called the IG tunnels.
Question: How could these tunnels be the product of evolutionary pressures, requiring long periods of time, if, in case the tunnel that protects the toxic intermediates is not instantiated from the beginning, the products would leak, and eventually kill the cell? This is an all-or-nothing business, where these tunnels had to be created right from the start, fully set up and developed.
These systems generally have the tunnel connecting two reactive centers, and the product of one reaction is transported to the second active site. In some cases, an IG tunnel network, instead of leading to another active site, can also lead to the lipid membrane so as to directly access the active site of membrane-bound enzymes. The substrate is generated within one of the active centers and is in the limiting amount as well as it could be toxic or unstable in the presented environment. Therefore, to ensure it reaches the destination reaction center, nature has devised strategies by constructing IG tunnels which, in several instances, are transient tunnels that only form upon entry of substates and have much more controlled and complex gating architectures. 57
Comment: This is truly fascinating evidence of intended design for important functions: To direct gases to where they are needed to perform a reaction.
Image description: The structure of carbamoyl phosphate synthetase
The small subunit that contains the active site for the hydrolysis of glutamine is shown in green. The N-terminal domain of the large subunit that contains the active site for the synthesis of carboxy phosphate and carbamate is shown in red. The C-terminal domain of the large subunit that contains the active site for the synthesis of carbamoyl phosphate is shown in blue. The two molecular tunnels for the translocation of ammonia and carbamate are shown in yellow dotted lines 56
Nucleotide metabolism: By evolution?
G. Caetano-Anollés (2013): The origin of metabolism has been linked to abiotic chemistries that existed in our planet at the beginning of life. While plausible chemical pathways have been proposed, including the synthesis of nucleobases, ribose and ribonucleotides, the cooption of these reactions by modern enzymes remains shrouded in mystery. Pathways of nucleotide biosynthesis, catabolism, and salvage originated ∼300 million years later byconcerted enzymatic recruitments and gradual replacement of abiotic chemistries. The simultaneous appearance of purine biosynthesis and the ribosome probably fulfilled the expanding matter-energy and processing needs of genomic information. 59
Comment: These are assertions, clearly not based on scientific data and observations, but ad-hoc conclusions that lack evidence.
56. Yubo Fan: A Combined Theoretical and Experimental Study of the Ammonia Tunnel in Carbamoyl Phosphate Synthetase 2009
57. Ruchi Anand: Tunnel Architectures in Enzyme Systems that Transport Gaseous Substrates December 3, 2021
58. Sérgio M. Marques: Role of tunnels, channels and gates in enzymatic catalysis 2016
59. Gustavo Caetano-Anollés: Structural Phylogenomics Reveals Gradual Evolutionary Replacement of Abiotic Chemistries by Protein Enzymes in Purine Metabolism March 13, 2013
 Copyright © 2022
Uncommon Descent
. This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.
Copyright © 2022
Uncommon Descent
. This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
November 8, 2022
At The Scientist: The Long and Winding Road to Eukaryotic Cells
Amanda Heidt writes:
This year, University of Paris-Saclay biologist Purificación López-García embarked with colleagues on a journey into life’s ancient past. The researchers traveled to the altiplanos of the northern Atacama Desert, high-altitude stretches of rocky soil and shrubbery in South America that are among the driest places in the world. Despite their inhospitable reputation, these plateaus may hold clues about the very origins of complex life. Amidst the dunes and barren mountains, there are pockets of life—warm, briny pools crusted over with colorful microbial mats of cyano-bacteria and archaea stacked atop one another like crepes. Long before Earth resembled its current state, López-García says, these microbial mats “were the forests of the past,” adding that scientists now use these clumps of microscopic life “as analogs of past ecosystems that certainly occurred at the time when eukaryotes first appear[ed].”
Each layer of these living mats is composed of different types of microbes that rely upon one another. At the surface, where light and oxygen are plentiful, photosynthesizing cyanobacteria dominate, while just below, heterotrophs that can persist in low-oxygen environments feed on their byproducts. Deeper down, the mats become dark and smelly, the result of the sulfate reducers and methanogens that populate these oxygen-bereft zones. Here, these partnerships become even more essential, with the castoffs of one group serving as fuel for another.
These close metabolic associations between organisms, a type of symbiosis known as syntrophy, may have prefaced the evolution of complex life by creating alliances that turned permanent over time, López-García says. In this way, individuals of different microbial species could have nested within one another to create a host with one or even several symbionts. This is exactly what scientists suspect happened to form a whole new type of cell, the eukaryote, which thrived and subsequently diversified into the macroscopic array of life we see today, including humans. So-called eukaryogenesis is not defined the same way by all researchers, but broadly, the term describes an evolutionary surge toward increasing cellular complexity between 1 and 2 billion years ago.
During this time, some of the defining characteristics of modern eukaryotic cells—the nucleus, mitochondria, cytoskeleton, cell membrane, and chloroplasts, among others—made their debut. These occurred between the first and last common ancestors of all living eukaryotes, known by their acronyms, FECA and LECA, respectively. Most of the details of these evolutionary leaps, however, remain unsettled. Researchers do not uniformly agree on which branch of life eukaryotes sprang from, which microbial players might have contributed to the process, or on the order of specific evolutionary milestones along the way. But the recent identification of the Asgard archaea, thought to be the closest living relatives to modern eukaryotes, has enlivened discussions about eukaryogenesis.
This year, University of Paris-Saclay biologist Purificación López-García embarked with colleagues on a journey into life’s ancient past. The researchers traveled to the altiplanos of the northern Atacama Desert, high-altitude stretches of rocky soil and shrubbery in South America that are among the driest places in the world. Despite their inhospitable reputation, these plateaus may hold clues about the very origins of complex life. Amidst the dunes and barren mountains, there are pockets of life—warm, briny pools crusted over with colorful microbial mats of cyano-bacteria and archaea stacked atop one another like crepes. Long before Earth resembled its current state, López-García says, these microbial mats “were the forests of the past,” adding that scientists now use these clumps of microscopic life “as analogs of past ecosystems that certainly occurred at the time when eukaryotes first appear[ed].

Each layer of these living mats is composed of different types of microbes that rely upon one another. At the surface, where light and oxygen are plentiful, photosynthesizing cyanobacteria dominate, while just below, heterotrophs that can persist in low-oxygen environments feed on their byproducts. Deeper down, the mats become dark and smelly, the result of the sulfate reducers and methanogens that populate these oxygen-bereft zones. Here, these partnerships become even more essential, with the castoffs of one group serving as fuel for another.
These close metabolic associations between organisms, a type of symbiosis known as syntrophy, may have prefaced the evolution of complex life by creating alliances that turned permanent over time, López-García says. In this way, individuals of different microbial species could have nested within one another to create a host with one or even several symbionts. This is exactly what scientists suspect happened to form a whole new type of cell, the eukaryote, which thrived and subsequently diversified into the macroscopic array of life we see today, including humans. So-called eukaryogenesis is not defined the same way by all researchers, but broadly, the term describes an evolutionary surge toward increasing cellular complexity between 1 and 2 billion years ago.
[Eukaryogenesis is] arguably one of the most important events in the history of life, after the origin of life itself.—Daniel Mills, Ludwig-Maximilians-Universität München
During this time, some of the defining characteristics of modern eukaryotic cells—the nucleus, mitochondria, cytoskeleton, cell membrane, and chloroplasts, among others—made their debut. These occurred between the first and last common ancestors of all living eukaryotes, known by their acronyms, FECA and LECA, respectively. Most of the details of these evolutionary leaps, however, remain unsettled. Researchers do not uniformly agree on which branch of life eukaryotes sprang from, which microbial players might have contributed to the process, or on the order of specific evolutionary milestones along the way. But the recent identification of the Asgard archaea, thought to be the closest living relatives to modern eukaryotes, has enlivened discussions about eukaryogenesis.
The Path to ComplexityToday, at the microbial mats in the Atacama Desert and other sites throughout the world, scientists are investigating what the earliest eukaryotic cells may have looked like, the partnerships they may have struck up with other organisms, and how their molecular machinery might have functioned and evolved. Already, the discovery of the Asgards has solidified certain aspects of eukaryogenesis while raising new questions about others. “I think this is the most exciting development in biology right now. So much is being discovered and so many predictions are being met,” says Daniel Mills, a geobiologist and postdoctoral researcher at Ludwig-Maximilians-Universität München who recently coauthored a paper suggesting that eukaryotes likely evolved in the absence of oxygen. Eukaryogenesis, he adds, is “arguably one of the most important events in the history of life, after the origin of life itself.”
Eukaryogenesis is broadly defined as the evolutionary path taken by increasingly complex lifeforms as they diverged from the simpler prokaryotes that dominated the early part of Earth’s biological history. The functional period of eukaryogenesis started just prior to the symbiosis between two prokaryotes and ended when the last common ancestor of modern eukaryotes arose. During this time, many of the most recognizable eukaryotic features appeared, including organelles such as mitochondria, nuclei, and chloroplasts, as well as cellular processes such as phagocytosis. The ordering of these events in time remains unclear.
 © NICOLLE FULLER, SAYO STUDIOORIGINAL HOST UNKNOWN
© NICOLLE FULLER, SAYO STUDIOORIGINAL HOST UNKNOWNORIGIN OF MITOCHONDRIAWhile the identity of original host in the symbiotic partnership that birthed modern eukaryotic cells remains mysterious, some researchers say the evidence suggests it was an archaeon rather than a bacterium. Scientists call this host, which lived more than a billion years ago, the first eukaryotic common ancestor, or FECA.
APPEARANCE OF UNIQUE FEATURESAt some point in the past, the prokaryote host formed a partnership with an alphaproteobacterium and permanently engulfed it, creating the mitochondrion. Researchers debate whether phagocytosis was needed to establish this relationship, but mitochondria did help power much of eukaryotes’ subsequent radiation.
BIRTH OF MODERN LIFENumerous other features and processes associated with modern eukaryotic cells evolved during this time, including the nucleus and cytoskeleton. The order of their appearance is uncertain.
The last eukaryotic ancestor (LECA) shared by all living eukaryotes today was already a complex cell by the time eukaryotes began to radiate. Over hundreds of millions of years, LECA gave rise to the complex organisms that exist today, including fungi, protists, plants, and animals.
See complete article at The Scientist.
“Mysterious…” It appears that evolutionary scientists are unable to offer any mechanism consistent with the known workings of nature that could explain eukaryogenesis. From my point of view as a physicist, just saying that unique features appeared, or that “ancestral” cells “gave rise to the complex organisms that exist today”, is mere story-telling. If someone tried to publish a physics paper postulating, for example, that superconductivity “arises from a low temperature environment”, and failed to give a mathematical or even a conceptual physics explanation for their postulate, it would justifiably never come print.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
At Evolution News: Does a New Scientific Study Offer Evidence of Life after Death?
Wesley J. Smith writes:
 Photo: Life after death, by Christopher Campbell, via Unsplash.
Photo: Life after death, by Christopher Campbell, via Unsplash.A new scientific study of people whose hearts stopped but received CPR and remained among us found that many remembered what are often called “near-death experiences.” From the NYU Grossman School of Medicine:
One in five people who survive cardiopulmonary resuscitation (CPR) after cardiac arrest may describe lucid experiences of death that occurred while they were seemingly unconscious and on the brink of death, a new study shows. . . .
Survivors reported having unique lucid experiences, including a perception of separation from the body, observing events without pain or distress, and a meaningful evaluation of life, including of their actions, intentions and thoughts toward others. The researchers found these experiences of death to be different from hallucinations, delusions, illusions, dreams or CPR-induced consciousness.
The study also detected some brain activity associated with the phenomenon:
A key finding was the discovery of spikes of brain activity, including so-called gamma, delta, theta, alpha and beta waves up to an hour into CPR. Some of these brain waves normally occur when people are conscious and performing higher mental functions, including thinking, memory retrieval, and conscious perception.
Life After Death?“These recalled experiences and brain wave changes may be the first signs of the so-called near-death experience, and we have captured them for the first time in a large study,” says Sam Parnia, MD, PhD, the lead study investigator and an intensive care physician, who is also an associate professor in the Department of Medicine at NYU Langone Health, as well as the organization’s director of critical care and resuscitation research. “Our results offer evidence that while on the brink of death and in a coma, people undergo a unique inner conscious experience, including awareness without distress.”
Does this prove that there is life after death? No. But I think it is evidence that could support that hypothesis. Indeed, that seems to be what the study authors conclude, at least between the lines:
No Natural-Selection Benefit
Identifying measureable electrical signs of lucid and heightened brain activity, together with similar stories of recalled death experiences, suggests that the human sense of self and consciousness, much like other biological body functions, may not stop completely around the time of death, adds Parnia.
“These lucid experiences cannot be considered a trick of a disordered or dying brain, but rather a unique human experience that emerges on the brink of death,” says Parnia. As the brain is shutting down, many of its natural braking systems are released. Known as disinhibition, this provides access to the depths of a person’s consciousness, including stored memories, thoughts from early childhood to death, and other aspects of reality. While no one knows the evolutionary purpose of this phenomenon, it clearly reveals “intriguing questions about human consciousness, even at death,” says Parnia.
Or, maybe there is no evolutionary explanation. There is certainly no discernible natural-selection benefit. Moreover, what purpose would such a “soft exit” offer? Why would it appear? How would it develop if consciousness is solely generated by the brain and is purely a materialistic phenomenon?
Perhaps these findings should be deemed circumstantial evidence of something beyond this existence, which moves us out of the scientific realm and into the theological. And — since those reporting on the experience claim to have engaged in an extensive life review — perhaps we should also consider the study as demonstrating that what we do and how we act in the here and now will impact our experience of whatever comes next.
If so, it would be prudent to govern ourselves accordingly, “for nothing is hidden that will not be made manifest, nor is anything secret that will not be known and come to light.”
Evolution News
Of course, the study referenced above is by no means the first scholarly investigation into near-death experiences (NDEs). If the testimony of thousands of people who have reported these experiences mean anything, there seems to be consistency with the concept of life after death and a continuing existence of the soul. An in-depth analysis of NDEs that I’ve found helpful is Imagine Heaven, by John Burke.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
November 6, 2022
At Nature: Dozens of papers co-authored by Nobel laureate raise concerns
Were we just now talking about the finding at Nature that a Nobel laureate’s papers were to get accepted?
So while we’re in this neck of the woods anyway, another Nature entry: “Concerns about image integrity have so far led to 17 retractions, corrections or expressions of concern for papers co-authored by geneticist Gregg Semenza.”
Commenters on the website PubPeer — where users scrutinize published research, often anonymously — have questioned images in 52 articles co-authored by Semenza that were published between 2000 and 2021. Since 2011, 17 of these papers have been retracted, corrected or had an expression of concern issued on them. The editorial notices cite the potential alteration, reuse or incorrect labelling of images showing experimental results. Another 15 of the papers are currently under investigation at their respective journals, Nature’s news team has learned.
Across the 32 papers that have so far drawn publisher scrutiny, all list Semenza as an author, but there are many combinations of different co-authors. – Holly Else
Is a pattern becoming apparent? Oh wait. There is no design in Nature, right? So they should quit complaining.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
At Nature: Authors’ names have “astonishing” influence in science
Why is this astonishing: “A Nobel prizewinner is six times more likely than someone less well known to get a thumbs-up for acceptance, finds study.”
Surely the author of this paywalled article, Nicola Jones, is joking if she claims to be astonished. Does she think we are all the slow class? Her mistake.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
George Ellis argues: We can’t know if the universe had a beginning
Cosmologist George Ellis at IAI News:
Most people today believe in The Big Bang theory when it comes to the origins of the cosmos. Can we be certain that the universe had a beginning? The history of the universe involves various stages. At very early times, it went through an extraordinarily rapid period of accelerating expansion when it became hugely bigger in a very short time; this is called inflation. At the end of inflation, that expansion had caused all the matter and radiation to dilute to almost zero, but then the field that had caused inflation decayed into very hot matter and radiation that continued expanding, but at a slower rate; that was the start of what we call the Hot Big Bang Era. The physical processes that occurred during this era are well understood, and all cosmologists agree on what happened then.
What we do not know is what happened before inflation began. The universe may or may not have had a beginning in that pre-inflationary era. The singularity theorems that Stephen Hawking developed do not apply, because the required energy conditions are now known to not be satisfied at that pre-inflationary time. In any case, a theory of quantum gravity is expected to apply at early enough times, but we don’t know what that theory is. To sum up: we do not know if the universe had a start, but we do know there was a Hot Big Bang.
Well, if the universe did not have a start, it must always have existed. And that’s the dreaded territory of Hilbert’s Hotel and infinity apart from mathematics.
You may also wish to read: Yes, you can manipulate infinity in math. The hyperreals are bigger (and smaller) than your average number — and better! (Jonathan Bartlett)
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
November 5, 2022
Supernovae and Black Holes
Aside from being the spectacular finale to a massive star’s life, what good is a supernova? Honestly, we wouldn’t be here without them. Both types of supernovae are nature’s mechanisms to both produce and disperse all of the heavier elements needed for life. As we have discussed, massive stars can produce elements via fusion burning with atomic numbers as high as that of iron (number 26), but if a supernova didn’t climax the process, all of those useful elements would forever be locked up deep within the stellar core. In addition, all of the elements heavier than iron (up to uranium, number 92) have no way of being produced in nature except during a star’s supernova explosion, when the vast amount of extra energy being released can be tapped to produce the heavier elements. These elements never form during the normal part of a star’s life, since their formation only proceeds by reactions that consume energy rather than producing it.
 Composite images of the Cas A supernova remnant, a structure resulting from the explosion of a star in the Cassiopeia constellation. Credits: X-ray: Chandra: NASA/CXC/SAO, IXPE: NASA/MSFC/J. Vink et al.; Optical: NASA/STScI.
Composite images of the Cas A supernova remnant, a structure resulting from the explosion of a star in the Cassiopeia constellation. Credits: X-ray: Chandra: NASA/CXC/SAO, IXPE: NASA/MSFC/J. Vink et al.; Optical: NASA/STScI.
Since the elements heavier than iron are only produced during the explosion phase of a supernova, we would expect that these elements are less abundant in our universe than the lighter elements, and such proves to be the case.1 Another prediction of the formation process of elements inside of stars, called stellar nucleosynthesis, is that the even-numbered elements (those with an even number of protons) will be more abundant than the odd-numbered elements. This also matches observational evidence, and the reason for it is that fusion most easily proceeds by adding a helium nucleus onto an already existing nucleus. The helium nucleus, with 2 protons and 2 neutrons, is like a brick in the process of element construction.
Throughout the history of a galaxy, supernovae serve to enrich the interstellar medium with the full array of elements needed for life. The preponderance of material in nebulae is still made up of just hydrogen and helium, but over time, about 2% of the total mix is built up to consist of elements heavier than helium. This enriched content of interstellar gases contains the right mixture to form not only a star like our sun, but also the variety of planets in our solar system, including ones with solid surfaces composed primarily of the heavier elements.
A Delicate Balance
Although supernovae serve a vital role in manufacturing life-essential elements, they also pose a serious danger to life. A supernova explosion releases prodigious amounts of radiation that would sterilize any planet orbiting around a nearby star system. Therefore, it’s crucial for us that the rate of supernovae in our galaxy has diminished throughout eons of time to a low level today—only 3 have been close enough to be observed with the naked eye in the last 1000 years, although others likely occurred further away. For this reason, our planet with its great variety of life could not have formed much earlier in the history of the galaxy than it did.2
Supernovae also play a role in determining where in the Galaxy the conditions are most ideal for a planet like Earth. Close in to the Galactic center, the density of stars in space is much higher than it is in our location (our sun is about two thirds of the way to the edge from the center of the galaxy). The higher density of stars near the central region of the Galaxy would increase the probability of sterilization events from supernova explosions. More crowded star conditions would also increase the risk of a gravitational encounter with another star, which could serve to disrupt our solar system’s planetary orbits. On the other hand, too far from the Galactic center the initial frequency of supernovae would have been too low to sufficiently enrich the interstellar medium with life-essential heavy elements. In actuality, our Solar System resides within a narrow band circling about 26,000 light years from the Galactic core, known to astronomers as the “Galactic Habitable Zone.”3 Earth’s location within our Galaxy’s habitable zone is ideal for allowing the long-term existence of life on our planet.
Black Holes
When a massive star reaches the end of its life and its iron core is crushed by gravity into a sphere of neutrons, if the mass of the resulting sphere is more than about twice the mass of our sun, gravity becomes so strong that the neutrons cannot withstand its crushing force. They then implode their mass into an even smaller volume, which pushes the escape velocity of the object past the speed of light, forming a black hole. Our current physics knowledge cannot adequately explain the properties of matter inside of a black hole, but perhaps that’s inconsequential, since no information from inside the black hole can ever reach the outside universe.
Black holes remind us of the delicate balance within the forces of nature. Although the universe needs the force of gravity to coalesce gases together in order to form galaxies and stars to give light, too much matter packed together in one place results in gravity forming a black hole from which no light emerges. Black holes also show us that our universe is not cyclical or eternal, for the formation of black holes is not reversible.
Black holes also spur us on to further understand the laws of physics of our universe, for although the General Theory of Relativity adequately describes and predicts the external properties of black holes, the conditions at the center of a black hole defy our current understanding of physics. Interestingly, the conditions at the beginning of our universe were similar to those at the center of a black hole, so a theory of physics which accurately described the center of a black hole would also help us to understand the first moments in the history of our universe. The needed theory would be a combination of quantum mechanics and gravity, accurately describing a state of matter with ultra-high mass in an ultra-small volume. Einstein sought to formulate such a theory, and many others have worked on it since his day, but so far to no avail. Undiscovered frontiers in science keep things interesting, and who knows when another under-employed “patent examiner” might surprise the world with the needed breakthrough.
1. Jeffrey Bennett, Megan Donahue, Nicholas Schneider, and Mark Voit, The Cosmic Perspective: The Solar System, 7th ed. (Pearson, San Francisco, 2014), p. 351.
2. Hugh Ross, Why the Universe is the Way it is (Baker Books, Grand Rapids, 2008) pp. 35, 177.
3. Guillermo Gonzalez, Donald Brownlee, and Peter Ward, “The Galactic Habitable Zone: Galactic Chemical Evolution,” Icarus 152 (July, 2001), pp. 185-200, cited in Hugh Ross, Why the Universe is the Way it is, p. 66.
Adapted from Canceled Science: What Some Atheists Don’t Want You to See, by Eric Hedin.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
At Science News: Astronomers have found the closest known black hole to Earth
Lisa Grossman writes:
The closest black hole yet found is just 1,560 light-years from Earth, a new study reports. The black hole, dubbed Gaia BH1, is about 10 times the mass of the sun and orbits a sunlike star.
 Stellar winds from a companion star might accrete onto black hole Gaia BH1 (illustrated), giving it a wispy halo distorted by gravity.GAIA/ESA, DPAC (CC BY-SA 3.0 IGO)
Stellar winds from a companion star might accrete onto black hole Gaia BH1 (illustrated), giving it a wispy halo distorted by gravity.GAIA/ESA, DPAC (CC BY-SA 3.0 IGO)
Most known black holes steal and eat gas from massive companion stars. That gas forms a disk around the black hole and glows brightly in X-rays. But hungry black holes are not the most common ones in our galaxy. Far more numerous are the tranquil black holes that are not mid-meal, which astronomers have dreamed of finding for decades. Previous claims of finding such black holes have so far not held up (SN: 5/6/20; SN: 3/11/22).
So astrophysicist Kareem El-Badry and colleagues turned to newly released data from the Gaia spacecraft, which precisely maps the positions of billions of stars (SN: 6/13/22). A star orbiting a black hole at a safe distance won’t get eaten, but it will be pulled back and forth by the black hole’s gravity. Astronomers can detect the star’s motion and deduce the black hole’s presence.
Out of hundreds of thousands of stars that looked like they were tugged by an unseen object, just one seemed like a good black hole candidate. Follow-up observations with other telescopes support the black hole idea, the team reports November 2 in Monthly Notices of the Royal Astronomical Society.
Gaia BH1 is the nearest black hole to Earth ever discovered — the next closest is around 3,200 light-years away. But it’s probably not the closest that exists, or even the closest we’ll ever find. Astronomers think there are about 100 million black holes in the Milky Way, but almost all of them are invisible. “They’re just isolated, so we can’t see them,” says El-Badry, of the Harvard-Smithsonian Center for Astrophysics in Cambridge, Mass.
The next data release from Gaia is due out in 2025, and El-Badry expects it to bring more black hole bounty. “We think there are probably a lot that are closer,” he says. “Just finding one … suggests there are a bunch more to be found.”
Science News
It’s quite an achievement to find a quiescent black hole, not only because it’s black, or not radiating, but because it’s so small. A 10 solar-mass black hole would have a radius (out to its event horizon) of only 30 km. Many asteroids in our solar system are bigger than this. The distance to Gaia BH1 is about the same as the distance to the Orion Nebula.
Copyright © 2022 Uncommon Descent . This Feed is for personal non-commercial use only. If you are not reading this material in your news aggregator, the site you are looking at is guilty of copyright infringement UNLESS EXPLICIT PERMISSION OTHERWISE HAS BEEN GIVEN. Please contact legal@uncommondescent.com so we can take legal action immediately.Plugin by Taragana
Michael J. Behe's Blog

- Michael J. Behe's profile
- 219 followers

